Иммунология
Т-лимфоциты: путешественники и домоседы
Природа. 2016. №2. С.8–14.
Касацкая С. А.

Иммунология
Т-лимфоциты: путешественники и домоседы
Клетки иммунной системы путешествуют по лимфе и кровотоку в поисках патогенов, распознают их и обезвреживают. Но значительная часть Т-лимфоцитов находится не в крови и не в лимфоузлах, а в органах, не относящихся к иммунной системе. Чем же заняты резидентные Т-клетки тканей, как они туда попадают и какие преимущества для медицины может дать их изучение?
 |
 |
|
|
Софья Алексеевна Касацкая |
Адекватная защитная реакция организма на вторжение вирусов, бактерий и других патогенов — уничтожить пораженные клетки, не допустив распространения инфекции и гибели большого числа собственных клеток. Если инфицированная вирусом клетка его заметила, запускаются процессы врожденного иммунитета: аутофагия (утилизация внутренних компонентов клетки с помощью ферментов лизосом) и апоптоз (запрограммированная гибель клеток). Однако патогенных вирусов и бактерий очень много, к тому же они постоянно меняются до неузнаваемости. Чтобы справиться с ними, подключается система адаптивного иммунитета и его главные участники — лимфоциты. Вершиной эволюции адаптивного иммунитета стал цитотоксический Т-лимфоцит, или Т-киллер. Для распознавания фрагмента вируса (антигена) на зараженной клетке он использует Т-клеточный рецептор (T cell receptor, TCR), случайно и независимо собирающийся на каждой Т-клетке в вилочковой железе (тимусе). Механизм сборки TCR уникален и присущ только иммунной системе позвоночных животных. Считается, что эти преимущества впервые получили примитивные рыбы около 500 млн лет назад, когда в результате ретровирусной инфекции в их гаметы внедрились гены, кодирующие особые белки (рекомбиназы), ответственные за рекомбинацию генов TCR.
Классическая иммунология человека построена на изучении иммунных клеток крови просто потому, что образец крови можно взять у любого пациента и исследовать в норме и в патологии. Именно на клетках крови была выстроена классификация Т-лимфоцитов — деление на Т-киллеры и Т-хелперы, которые проверяют антигенную специфичность Т-киллеров, выдают им «лицензию на убийство» и способны управлять всем ходом иммунного ответа через сигнальные растворимые молекулы, цитокины. Позднее из ветви Т-хелперов была выделена группа регуляторных Т-клеток, подавляющих избыточный адаптивный иммунитет.
Но, как нам напоминает реклама йогурта, значительная часть клеток иммунной системы сосредоточена вокруг слизистой оболочки пищеварительного тракта и в других тканях. В то время как в 5 – 6 л крови взрослого человека находится около 6 – 15 млрд T-лимфоцитов, в эпидермисе и коже — 20 млрд Т-клеток [1]Clark R.A. Skin resident T cells: the ups and downs of on site immunity // J. Invest. Dermatol. 2010. V.130. P.362–370.Doi:10.1038/jid.2009.247 в печени — еще 4 млрд [2]Doherty D.G., Norris S., Madrigal-Estebas L. et al. The human liver contains multiple populations of NK cells, T cells, and CD3+CD56+ natural T cells with distinct cytotoxic activities and Th1, Th2, and Th0 cytokine secretion patterns // J. Immunol. 1999. V.163. P.2314–2321. Достаточно ли изучения образцов крови для полного описания функций Т-клеток, если в периферических органах Т-клеток больше, чем в кровотоке? И достаточно ли классических субпопуляций, чтобы описать все типы Т-клеток, находящихся в организме человека?
Жизненный цикл Т-лимфоцита
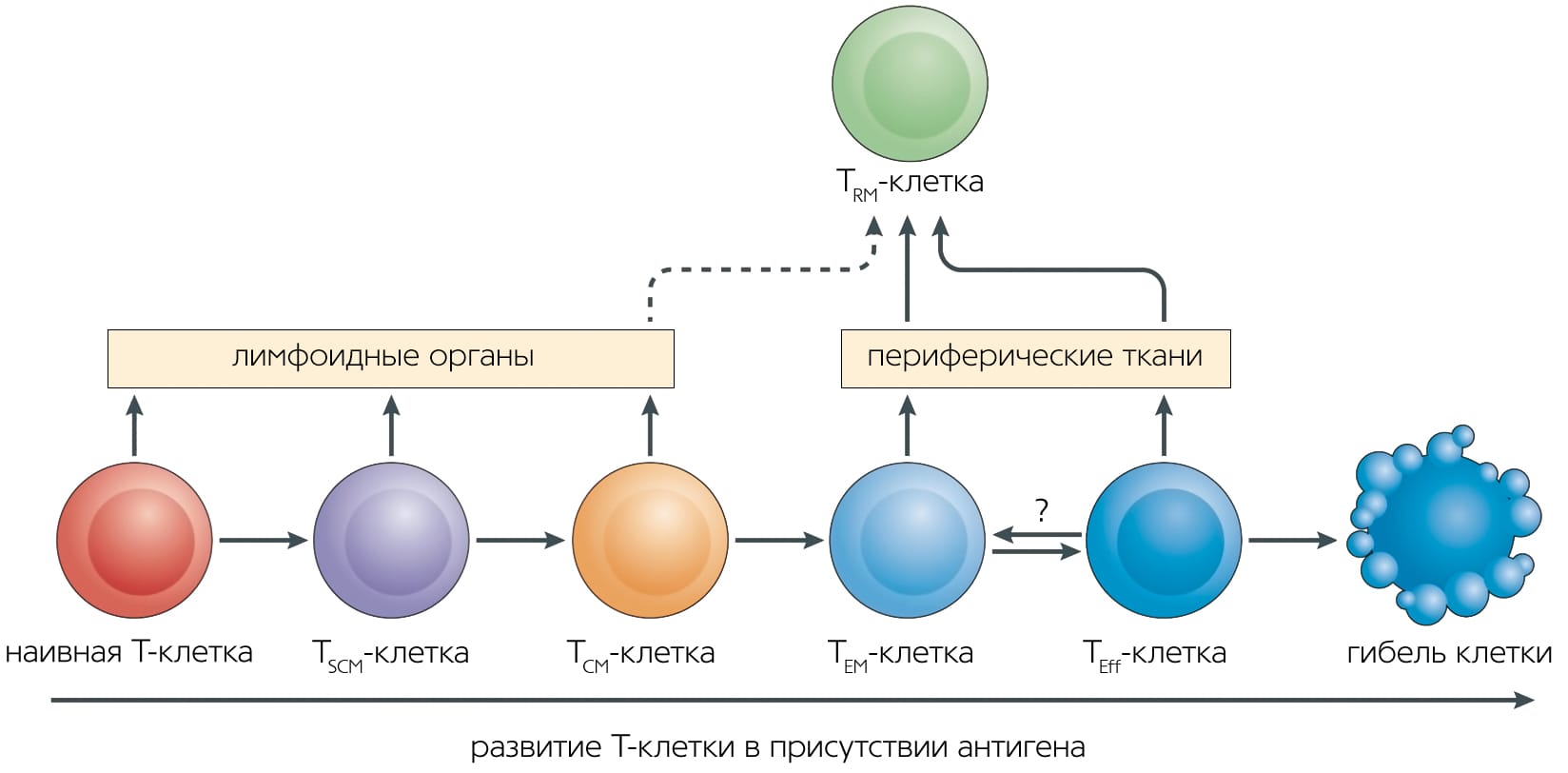
Каждая Т-клетка после сборки TCR проходит тестирование на функциональность случайно собранного рецептора (положительная селекция) и на отсутствие специфичности к собственным антигенам организма, т.е. на отсутствие очевидной аутоиммунной угрозы (отрицательная селекция). Этапы селекции происходят в вилочковой железе; при этом более 90% клеток-предшественников погибает, не сумев правильно собрать рецептор либо пройти селективный отбор. Выжившие Т-клетки пролиферируют и выходят из тимуса в кровоток — это наивные Т-лимфоциты, еще не встречавшиеся с антигеном. Наивная Т-клетка циркулирует в крови и периодически заходит в лимфоузлы, где в Т-клеточной зоне контактирует со специализированными клетками, которые представляют ей чужеродный антиген.
После встречи с антигеном в лимфоузле Т-клетка приобретает способность снова делиться — становится предшественницей Т-клеток памяти (Stem Cell Memory T cells, TSCM). Cреди ее потомков появляются клетки центральной памяти (Central Memory T cells, TCM) и эффекторные клетки-предшественники (Effector Memory T cells, TEM), которые при делении дают короткоживущие эффекторные клетки, осуществляющие иммунную реакцию (TEMRA-клетки) [3]Iijima N., Iwasaki A. Tissue instruction for migration and retention of TRM cells // Trends Immunol. 2015. V.36. P.556–564.Doi:10.1016/j.it.2015.07.002 Все эти клетки выходят из лимфоузла и перемещаются по крови. Эффекторные клетки затем могут покинуть кровоток для осуществления иммунной реакции в периферической ткани органа, где находится патоген. Что потом — снова путешествие по крови и лимфоузлам?
Клетки стромы, т.е. основы лимфоузла, выделяют сигнальные вещества (хемокины) для того, чтобы позвать Т-клетку в лимфоузел. Распознают хемокины лимфоузлов рецепторы хоминга CCR7 и CD62L. Но на эффекторных клетках оба рецептора отсутствуют. Из-за этого долгое время было загадкой, как эффекторные клетки могут попасть из периферической ткани обратно во вторичные лимфоидные органы — селезенку и лимфоузлы.
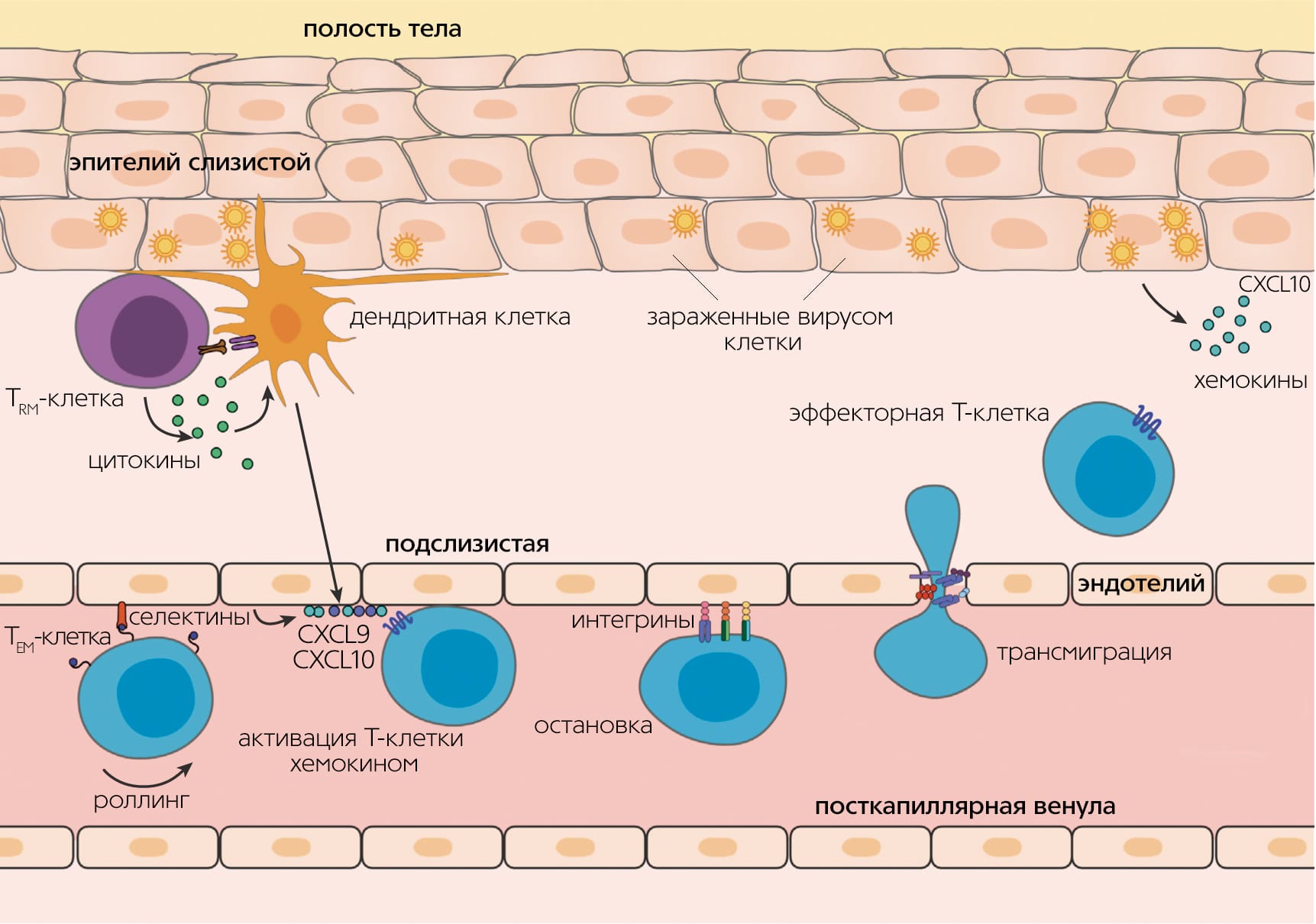
Трансмиграция лимфоцитов. Видео: www.xvivo.net
В то же время стали накапливаться данные (о различиях в репертуарах TCR и профилях транскрипции между TEM в крови и в других тканях), которые никак не укладывались в концепцию постоянной миграции Т-клеток между тканями и кровью. Решено было выделить новую субпопуляцию — резидентные клетки памяти (Resident Memory T cells, TRM), которые населяют определенный орган и не рециркулируют [5]Schenkel J.M., Masopust D. Tissue-resident memory T cells // Immunity. 2014. V.41. P.885–897.Doi:10.1016/j.immuni.2014.12.007
Откуда впервые появляются резидентные клетки ткани? Это потомки эффекторных клеток, которые потеряли способность рециркулировать. Некоторые периферические для иммунной системы ткани, например слизистая тонкого кишечника и брюшная полость, позволяют эффекторным Т-лимфоцитам проникать внутрь свободно, другие — очень ограниченно. Большой поток эффекторных Т-клеток в эти ткани наблюдается только при реакции воспаления. К тканям второго типа относятся головной и спинной мозг, отделенные барьером от иммунной системы, а также многие другие ткани: периферические ганглии, слизистые половых органов и кишечника, легкие, эпидермис, глаза. Разница между двумя типами тканей — в экспрессии дополнительных молекул хоминга для эффекторных Т-клеток, например молекул адгезии MadCAM-1 для проникновения в эпителий [3]Iijima N., Iwasaki A. Tissue instruction for migration and retention of TRM cells // Trends Immunol. 2015. V.36. P.556–564.Doi:10.1016/j.it.2015.07.002
Резидентные Т-клетки в старении тканей человека
Карта соотношений присутствия отдельных субпопуляций Т-клеток в разных органах человека, как ни странно, была составлена только в 2014 г. Команда Донны Фарбер из медицинского центра Колумбийского университета Нью-Йорка провела сравнение фенотипов Т-клеток, выделенных из крови и тканей доноров органов всех возрастных групп от 3 до 73 лет (всего 56 доноров) [6]Thome J.J., Yudanin N., Ohmura Y. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life // Cell. 2014. V.159. P.814–828.Doi:10.1016/j.cell.2014.10.026 Анализ субпопуляций Т-клеток при помощи проточной цитофлуориметрии подтвердил многие данные, полученные методами с меньшим разрешением и меньшей статистикой, и некоторые черты описания иммунной системы, перенесенные с иммунологии мыши на человека, к примеру снижение содержания наивных Т-лимфоцитов во всех органах при старении организма.
Уменьшение числа наивных Т-клеток с возрастом связано с быстрым старением вилочковой железы, в которой будущие Т-клетки проходят этапы сборки TCR, проверку его работоспособности и селекцию на отсутствие аутоиммунного потенциала. Важно не только снижение абсолютной численности наивных Т-клеток, но и уменьшение разнообразия репертуара Т-клеточных рецепторов, а значит, и возможности сформировать адаптивный иммунный ответ на ранее незнакомую инфекцию [7]Britanova O.V., Putintseva E.V., Shugay M. et al. Age-related decrease in TCR repertoire diversity measured with deep and normalized sequence profiling // J. Immunol. 2014. V.192. P.2689–2698.Doi:10.4049/jimmunol.1302064 Для наивных Т-киллеров подтвердилось прогрессирующее падение численности в крови и лимфоузлах, хотя для наивных Т-хелперов отрицательная корреляция численности с возрастом в данном исследовании оказалась значительной только для вторичных лимфоидных органов, но не для крови.
Выделение Т-лимфоцитов памяти, эффекторных клеток памяти и короткоживущих эффекторных клеток из слизистых легких, тонкого и толстого кишечника, паховых и мезентериальных лимфоузлов доноров органов позволило впервые оценить динамику данных популяций в тканях человека при старении. Доля центральных клеток памяти ожидаемо растет с течением жизни, в соответствии с ростом числа инфекций, которые успели встретиться организму и попасть в библиотеку памяти иммунной системы. Эффекторные клетки памяти TEM стремительно заполняют нишу для Т-клеток в тканях ребенка, быстро, примерно к 12 годам, вытесняя наивные Т-клетки. Растет и количество терминально дифференцированных эффекторных Т-киллеров (TEMRA), но только в лимфоузлах и в селезенке; в нелимфоидных тканях численность TEMRA падает. Короткоживущие терминально дифференцированные Т-киллеры чаще всего встречаются в крови, селезенке и слизистых легких в любом возрасте, а вот среди Т-хелперов эта субпопуляция представлена исчезающе малым числом клеток. Аналогично мало центральных клеток памяти среди Т-киллеров, преимущественно они находятся в слизистых двух барьерных тканей: легких и кишечника.
Широкими мазками карту распределения Т-лимфоцитов человека можно обрисовать так: наивные Т-клетки путешествуют по крови и периодически заходят во вторичные лимфоидные органы, киллеры TEMRA находятся в крови, селезенке и легких. Для центральных клеток памяти, судя по всему, характерно более индивидуальное распределение по тканям, чем для других субпопуляций: во всяком случае, закономерностей динамики при старении разных тканей выявить не удалось. Эффекторные клетки памяти, включающие TRM-субпопуляцию, доминируют среди Т-клеток слизистых барьерных тканей. В целом, при старении Т-клеточного иммунитета нелимфоидные ткани проявляют большую стабильность субпопуляций, лимфоидные ткани — большую возрастную динамику типов Т-клеток [6]Thome J.J., Yudanin N., Ohmura Y. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life // Cell. 2014. V.159. P.814–828.Doi:10.1016/j.cell.2014.10.026 Стабильность тканевых клеток проще объяснить, если разобраться, какие из эффекторных клеток TEM остаются в ткани, становятся резидентными TRM и из каких событий состоит их жизнь после отказа от путешествий по организму.

Как отличить резидентные клетки тканей от примесей клеток крови?
Резидентные Т-клетки корректно, но неудобно каждый раз определять по способности индивидуальной клетки мигрировать в лимфоузлы, поэтому необходимо составить список характерных признаков, по которым можно выявить принадлежность к этой субпопуляции. Резидентные Т-лимфоциты в тканях — естественных барьерах организма (например в легких и слизистой тонкого кишечника) немного похожи на классические эффекторные клетки крови: экспрессируют маркер активированных клеток CD69, причем экспрессия стабильна в течение жизни при взрослении и старении и характерна для всех нелимфоидных тканей. Но вдобавок CD69 колокализуется с маркером CD103, который обозначает группу молекул адгезии — интегринов, способствующих прикреплению резидентной Т-клетки к эпителию и к фибробластам в подслизистой выбранного органа. Для эффекторных Т-клеток во вторичных лимфоидных органах экспрессия интегринов CD103 совершенно нехарактерна: TEM-клетки постоянно сохраняют подвижный фенотип.
У карты, составленной коллективом Донны Фарбер, есть крупный недочет: неясно, насколько чисто удается выделить Т-лимфоциты из органа, какую долю анализируемых клеток на самом деле составляют Т-лимфоциты крови из капилляров внутри органа.
Особенно остро вопрос загрязнения клетками крови стоит для легких — неслучайно субпопуляционный состав Т-клеток легких неожиданно похож на Т-клетки крови и лимфоузлов. Вопрос загрязнения клетками крови был изящно решен для Т-лимфоцитов мыши: подопытных животных заражали вирусом лимфоцитарного хориоменингита после пересадки трансгенного клона Т-клеток P14, специфичного к данному вирусу. В результате при инфекции большая часть циркулирующих клеток была представлена вирусоспецифичным клоном P14, а его присутствие в тканях можно было выявить с помощью флуоресцирующих антител к TCR P14. Мышам в кровь вводили антитело анти-CD8 к маркеру Т-киллерных клеток, оно быстро распространялось по кровотоку и связывалось со всеми Т-киллерами в крови (но не в тканях). При микроскопии срезов органов легко было отличить резидентные киллерные TRM от только недавно вышедших из крови в орган клеток, помеченных анти-CD8 антителом [9]Steinert E.M., Schenkel J.M., Fraser K.A. et al. Quantifying memory CD8 T cells reveals regionalization of immunosurveillance // Cell. 2015. V.161. P.737–749.Doi:10.1016/j.cell.2015.03.031 Численность резидентных клеток, подсчитанная этим методом, в 70 раз превышала количество, определенное методом проточной цитометрии; разница меньше чем в два раза наблюдалась только для резидентных клеток лимфоузлов и селезенки. Получается, стандартные методики выделения лимфоцитов из органов плохо подходят для анализа киллерных резидентных клеток и существенно занижают размеры популяции.
Работа резидентных Т-клеток: не стоит путать туризм с эмиграцией
Мышиные резидентные клетки тканей в нормальной ситуации почти не перемещаются внутри нелимфоидной ткани и достаточно прочно прикреплены молекулами адгезии к строме органа. Когда резидентные макрофаги той же ткани секрецией цитокинов инициируют реакцию воспаления, TRM приобретают большую подвижность и патрулируют близлежащий эпителий в поисках зараженных клеток.
Если воспалительная реакция усиливается, то клетки воспринимают это как сигнал о подкреплении: к работе патрульных TRM подключаются вновь прибывающие из крови TCM и TEM. Эти клетки крови куда более подвижны и лучше перемещаются в эпителии. Значит ли это, что именно в крови находятся готовые действовать Т-киллеры среди TEM, а CD8+TRM выполняют в ткани хелперные и регуляторные функции?
C одной стороны, Т-хелперы по спектру Т-клеточных рецепторов более тканеспецифичны, т.е. пересечений между репертуарами TCR клеток, взятых из разных тканей, совсем мало, тогда как клетки одного клона Т-киллера встречаются в разных тканях среди TEM [6]Thome J.J., Yudanin N., Ohmura Y. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life // Cell. 2014. V.159. P.814–828.Doi:10.1016/j.cell.2014.10.026 Спектр функций и репертуар антигенной специфичности TRM еще предстоит исследовать, но способности к уничтожению зараженных клеток тканей у TRM-киллеров точно есть. Более того, в модели мышиной инфекции полиомавирусом, протекающей в ткани головного мозга, аффинность вирусо-специфичных Т-клеточных рецепторов резидентных киллерных клеток выше, чем у вирусоспецифичных клеток центральной памяти [10]Frost E.L., Kersh A.E., Evavold B.D., Lukacher A.E. Cutting edge: resident memory CD8 T cells express high- affinity TCRs // J. Immunol. 2015. V.195. P.3520–3524.Doi:10.4049/jimmunol.1501521
Однако размер популяции Т-клеток зависит не только от специфичности TCR к инфекциям, которые раньше протекали в данном органе, но и от гомеостатической пролиферации Т-клеток — размножения более удачливых клеток для заполнения емкости органа по числу Т-лимфоцитов. По маркерам CD28 и CD127 на поверхности клеток можно отличить недавно и давно активированные через TCR клетки от тех, которые получили только гомеостатический сигнал к пролиферации от фактора роста IL-7. При старении ткани гомеостатическое размножение клеток начинает преобладать над пролиферацией активированных через TCR-клеток.
Независимо от Т-клеточных рецепторов часто функционируют NKT-клетки, тип резидентных клеток печени, встречающихся и в других тканях. Они могут быть активированы NK-клеточными рецепторами через распознавание не индивидуальных антигенов, а общих молекулярных паттернов опасности и тканевого стресса. При активации CD8+ NKT-клетки выделяют цитотоксические гранулы и лизируют подозрительные клетки ткани, к примеру единичные опухолевые клетки и зараженные вирусами клетки, экспрессирующие и выставляющие на внешней мембране стрессорные молекулы. При старении тенденция TRM к активации без Т-клеточного рецептора, через NK-клеточные рецепторы или цитокиновые сигналы, может приводить к ошибочному лизису клеток ткани, недостаточному контролю над хронически зараженными или перерождающимися участками эпителия.
Патологические проявления, связанные с работой резидентных Т-клеток, включают органоспецифичные аутоиммунные синдромы и синдромы хронического воспаления в ткани. Примеры хронического воспаления, поддерживаемого резидентными Т-лимфоцитами, — контактный дерматит и псориаз, а механизмом служит выделение воспалительных факторов IL-17 резидентными Т-киллерами и IL-22 резидентными Т-хелперами дермы. CD8+ эффекторные Т-киллеры, находящиеся в головном мозге, похожи по совокупности мембранных молекул-маркеров TRM кожи, кишечника и легких и способны подталкивать развитие перемежающегося рассеянного склероза при периодических выбросах воспалительных цитокинов. Неясно, однако, есть ли в норме в головном мозге популяция TRM или же это Т-лимфоциты, оставшиеся в ткани после нейротропной вирусной инфекции [8]Park C.O., Kupper T.S. The emerging role of resident memory T cells in protective immunity and inflammatory disease // Nat. Med. 2015. V.21. P.688–697.Doi:10.1038/nm.3883
Функции резидентных клеток памяти в норме — при отсутствии инфекции или хронического воспаления — могут включать cross-talk (взаимную регуляцию преимущественно через секрецию цитокинов и костимуляторные молекулы) с неклассическими малоизученными лимфоидными клетками. Ими могут быть ассоциированные со слизистыми γ/δ Т-клетки, несущие альтернативный вариант сборки Т-клеточного рецептора, или лимфоидные клетки врожденного иммунитета (Innate Lymphoid Cells, ILC), которые делят с Т- и В-лимфоцитами общие черты эпигенетического ландшафта, но не имеют Т-/В- или NK-клеточныхрецепторов [11]Diverse functions of mucosal resident memory T cells / Eds K.S.Schluns, K.Klonowski // Frontiers in Immunology [Electronic resource]. N.Y., 2015.Doi:10.3389/fimmu.2015.00090[12]Godfrey D.I., Uldrich A.P., McCluskey J. et al. The burgeoning family of unconventional T cells // Nat. Immunol. 2015. V.16. P.1114–1123.Doi:10.1038/ni.3298
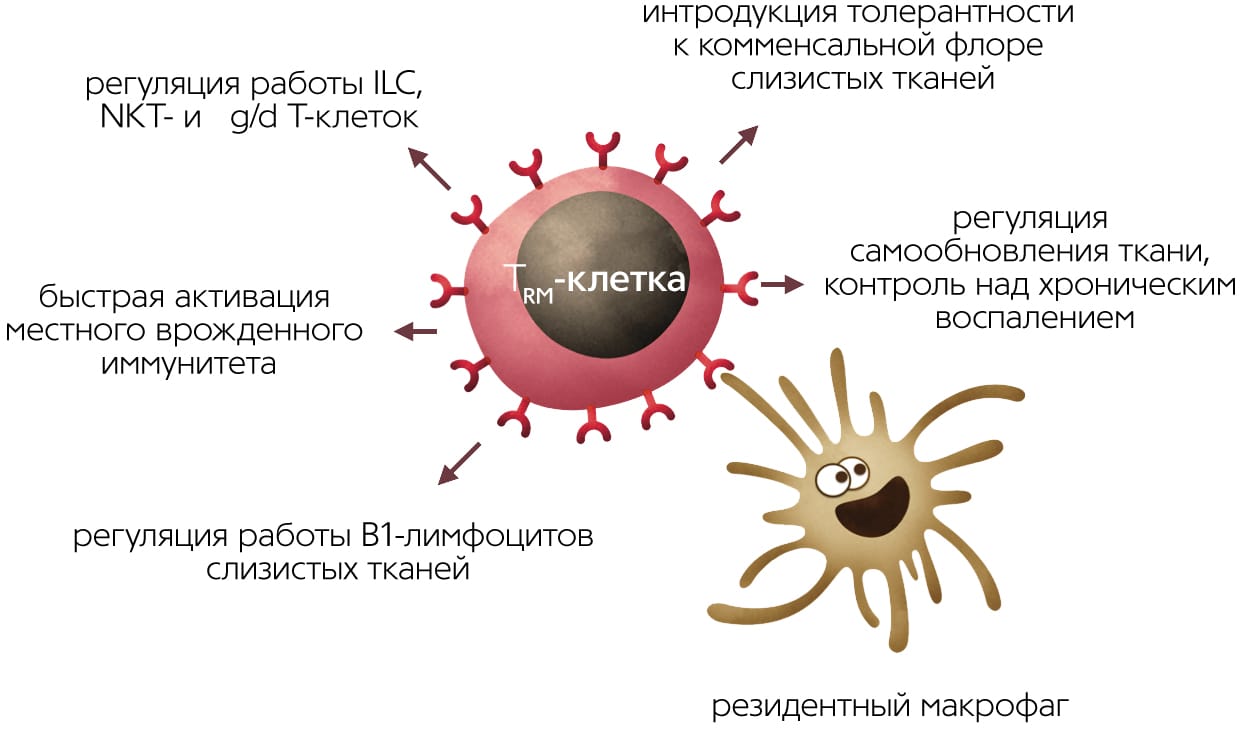
TRM-клетки контактируют с антигенпрезентирующими клетками тканей — дендритными клетками кожи и резидентными макрофагами тканей. Резидентные миелоидные клетки в разных тканях дифференцированы и слабо похожи друг на друга. К примеру, макрофаги маргинальной зоны селезенки, макрофаги печени и микроглия (макрофаги мозга) будут сильно отличаться и по морфологии, и по спектру функций. Кроме обнаружения антигенов в ткани, резидентные макрофаги заняты регуляцией процессов старения и самообновления тканей, в частности, выделяют факторы роста и цитокины, стимулирующие деление стволовых клеток тканей. В жировой ткани, к примеру, макрофаги стимулируют дифференцировку новых жировых клеток, но при переходе в активированное M1-состояние запускают воспаление и вместо дифференцировки заставляют увеличиваться и набухать имеющиеся жировые клетки. Сопутствующие изменения метаболизма жировой ткани приводят к накоплению жировой массы и в последние годы связываются с механизмами развития ожирения и диабета II типа. В коже цитокины, выделяемые макрофагами и резидентными γ/δ Т-клетками, стимулируют деление стволовых клеток при регенерации эпидермиса и стволовых клеток волосяных фолликулов [13]Castellana D., Paus R., Perez-Moreno M. Macrophages contribute to the cyclic activation of adult hair follicle stem cells // PLoS Biol [Electronic resource]. 2014. V.12. e1002002.Doi:10.1371/journal.pbio.1002002[14]Rodero M.P., Khosrotehrani K. Skin wound healing modulation by macrophages // Int. J. Clin. Exp. Pathol. 2010. V.3. №7. P.643–653. Можно предположить, что хелперные TRM-клетки при патрулировании эпителия и образовании контактов с тканевыми макрофагами могут модулировать спектр и объем выделяемых последними факторов роста для стволовых клеток, воспалительных цитокинов и факторов ремоделирования эпителия — и тем самым участвовать в обновлении тканей.
Что изучение TRM может дать медицине?
Понимание принципов работы резидентных Т-клеток абсолютно необходимо для борьбы с инфекциями, которые не поступают сразу в кровь, а проникают в организм через барьерные ткани, т.е. для подавляющего большинства инфекций. Рациональный дизайн вакцин для защиты от этой группы инфекций может быть направлен именно на усиление первого этапа защиты с помощью резидентных клеток. Ситуация, при которой оптимально активированные специфичные к антигену клетки элиминируют патоген в барьерной ткани, куда выгоднее, чем запуск острого воспаления для вызова Т-лимфоцитов из крови, поскольку меньше повреждается ткань.
Репертуар TCR, ассоциированных со слизистыми барьерных тканей, считается частично вырожденным и наиболее распространенным, т.е. идентичным для многих людей в популяции. Тем не менее искажения при выделении Т-клеток из органов, перекос данных в результате отбора в когорты только определенных европеоидных доноров и общее небольшое количество накопленных данных секвенирования не дают уверенности в публичности репертуаров Т-клеточных рецепторов TRM-клеток. Впрочем, это было бы удобно: дизайн вакцин мог бы сводиться к поиску и модификации наиболее аффинных и иммуногенных пептидов в патогене, взаимодействующих с одним из распространенных вариантов TCR в барьерной для этого патогена ткани.
Конечно, представления о том, какие TCR несут на своей поверхности TRM-клетки, недостаточно для того, чтобы эффективно манипулировать иммунными реакциями в ткани. Предстоит детально изучить факторы, влияющие на заселение тканей определенными клонами Т-клеток, и разобраться в механизмах активации местного тканевого иммунитета и индукции толерантности TRM. Как заселяются ниши Т-лимфоцитов в слизистых у ребенка до встречи с большим числом патогенов и, соответственно, до формирования значительного пула эффекторных Т-клеток памяти — предшественников резидентных клеток и клеток центральной памяти? Почему и как вместо классической активации лимфоцитов формируется реакция толерантности к микробам непатогенной флоры слизистых? Эти вопросы стоят на повестке дня в изучении резидентных клеток иммунной системы.
Определение закономерностей хоминга Т-лимфоцитов в определенные ткани может дать преимущество в клеточной иммунотерапии опухолевых заболеваний. Теоретически киллерные Т-клетки нужной специфичности к опухолевому антигену, активированные in vitro, должны убивать опухолевые клетки пациента. На практике подобная иммунотерапия осложняется тем, что опухолевые клетки способны подавлять иммунные реакции и приводить в неактивное состояние приближающиеся к опухоли Т-киллеры. Зачастую в массе растущей опухоли и вокруг нее накапливаются анергичные Т-лимфоциты — в первую очередь TRM данной ткани. Из множества инъецированных пациенту активных опухолеспецифичных Т-клеток до цели дойдут немногие, и даже они могут оказаться практически бесполезными в иммуносупрессивном микроокружении опухоли.
Расшифровка механизмов, которые обеспечивают попадание конкретных клонов Т-клеток в определенные ткани, может позволить более эффективно направлять к опухоли сконструированные в лаборатории Т-лимфоциты и приблизить эру доступной персонализированной иммунотерапии.
Литература
- Clark R.A. Skin resident T cells: the ups and downs of on site immunity // J. Invest. Dermatol. 2010. V.130. P.362– 370.
- Doi:10.1038/jid.2009.247
- Doherty D.G., Norris S., Madrigal-Estebas L. et al. The human liver contains multiple populations of NK cells, T cells, and CD3+CD56+ natural T cells with distinct cytotoxic activities and Th1, Th2, and Th0 cytokine secretion patterns // J. Immunol. 1999. V.163. P.2314–2321.
- Iijima N., Iwasaki A. Tissue instruction for migration and retention of TRM cells // Trends Immunol. 2015. V.36. P.556–564.
- Doi:10.1016/j.it.2015.07.002
- Farber D., Yudanin N., Restifo N.P. Human memory T cells: generation, compartmentalization and homeostasis // Nat. Rev. Immunol. 2014. V.14. P.24–35.
- Doi:10.1038/nri3567
- Schenkel J.M., Masopust D. Tissue-resident memory T cells // Immunity. 2014. V.41. P.885–897.
- Doi:10.1016/j.immuni.2014.12.007
- Thome J.J., Yudanin N., Ohmura Y. et al. Spatial map of human T cell compartmentalization and maintenance over decades of life // Cell. 2014. V.159. P.814–828.
- Doi:10.1016/j.cell.2014.10.026
- Britanova O.V., Putintseva E.V., Shugay M. et al. Age-related decrease in TCR repertoire diversity measured with deep and normalized sequence profiling // J. Immunol. 2014. V.192. P.2689–2698.
- Doi:10.4049/jimmunol.1302064
- Park C.O., Kupper T.S. The emerging role of resident memory T cells in protective immunity and inflammatory disease // Nat. Med. 2015. V.21. P.688–697.
- Doi:10.1038/nm.3883
- Steinert E.M., Schenkel J.M., Fraser K.A. et al. Quantifying memory CD8 T cells reveals regionalization of immunosurveillance // Cell. 2015. V.161. P.737–749.
- Doi:10.1016/j.cell.2015.03.031
- Frost E.L., Kersh A.E., Evavold B.D., Lukacher A.E. Cutting edge: resident memory CD8 T cells express high- affinity TCRs // J. Immunol. 2015. V.195. P.3520–3524.
- Doi:10.4049/jimmunol.1501521
- Diverse functions of mucosal resident memory T cells / Eds K.S.Schluns, K.Klonowski // Frontiers in Immunology [Electronic resource]. N.Y., 2015.
- Doi:10.3389/fimmu.2015.00090
- Godfrey D.I., Uldrich A.P., McCluskey J. et al. The burgeoning family of unconventional T cells // Nat. Immunol. 2015. V.16. P.1114–1123.
- Doi:10.1038/ni.3298
- Castellana D., Paus R., Perez-Moreno M. Macrophages contribute to the cyclic activation of adult hair follicle stem cells // PLoS Biol [Electronic resource]. 2014. V.12. e1002002.
- Doi:10.1371/journal.pbio.1002002
- Rodero M.P., Khosrotehrani K. Skin wound healing modulation by macrophages // Int. J. Clin. Exp. Pathol. 2010. V.3. №7. P.643–653.